Миелинизация нервных волокон спинного мозга. Миелинизация нервных волокон зрительного пути
Отдельных нейронов обычно объединяются в пучки – нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 2.3). Чехол прерывается примерно через каждые 0,5-1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис. 2.9 показано, как это происходит. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном это происходит за счет клеток олигодендроглии.
Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2 / 3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга.
Известно, что у новорожденного ребенка миелинизировано примерно 2 / 3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция , он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится . Это заболевание развивается незаметно и очень медленно, последствием является паралич движения.
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о и температуре.
Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис. 2.10 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы,-выходящий ток, который деполяризует соседний невозбужденный участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность – зона невозбудимости. Нам уже известно, что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается . Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100 – 120 м/с, должны иметь диаметр около 1000 мкм (1 мм).
У млекопитающих животных природа сохранила немиелинизированными только те возбуждение о боли, температуре, управляют медленно работающими внутренними органами мочевым волокна, которые проводят органами – мочевым пузырем, кишечником и пр. Практически все нервные волокна в человека имеют миелиновые чехлы. На рис. 2.11 показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые находятся на стыке между двумя участками миелина. Напомним, что каждый участок образован только одной клеткой, поэтому это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. Мембрана аксона между двумя соседними миелиновыми чехлами оказывается не покрытой миелином (так называемый перехват Ранвье). Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату. Это похоже на те волшебные сапоги-скороходы, которые надевал кот в известной сказке, мгновенно переносясь из одного места в другое.
Челябинская государственная медицинская академия
Кафедра гистологии, цитологии и эмбриологии
Лекция
«Нервная ткань. Нервные волокна и нервные окончания»
2003 г.
План
1. Понятие о нервном волокне
2. Характеристика безмиелиновых нервных волокон.
3. Характеристика миелиновых нервных волокон.
4. Периферический нерв: понятие, строение, оболочки, регенерация.
5. Синапсы: понятие, классификации по локализации, эффекту, эволюции, характеру нейротрансмиттера, строению.
6.Нервные окончания: понятия, разновидности, строение чувствительных и двигательных нервных окончаний.
Список слайдов
1.Тельце Фатер- Пачини 488.
2.Миелиновын нервные волокна 446
3.Поперечный срез периферического нерва 777.
4.Нервные синапсы на поверхности мультиполярной нервной клетки 789.
5.Тельце Фатер- Пачини и тельце Мейснера 784.
6.Тельце Мейснера 491.
7.Тельце Мейснера 786.
8.Свободные нервные окончания в эпителии
9.Свободные нервные окончания в эпидермисе 782.
10.Двигательные нервные окончания в скелетной мышце 785.
11.Синапс (схема) 778.
12.Ультраструктура синапсов 788
13.Миелиновые нервные волокна 780
14.Безмиелиновые нервные волокна 444.
15.Миелинизация нервных волокон 793.
16.Нервный пучок 462.
17.Нервнол-мышечное окончание 487.
18.Инкапсулированные нервные окончания 450.
Нейроны, лежащие в центральной нервной системе и в ганглиях, связаны с периферией при помощи своих отростков: дендритов и аксонов. Выходя на периферию, отростки нервных клеток покрываются оболочками, в результате чего образуются нервные волокна. Каждое нервное волокно содержит, таким образом, отросток нервной клетки (аксон или дендрит)- осевой цилиндр и оболочку, построенную из клеток глии- глиальную оболочку. По строению глиальной оболочки различают миелиновые (мякотные) нервные волокна и безмиелиновые (безмякотные) нервные волокна.
Безмиелиновые (безмякотные) нервные волокна преимущественно встречаются в вегетативной нервной системе. Вырастающие отростки нервных клеток покрываются клетками олигодендроглии, которые принято называть в периферической нервной системе шванновскими клетками или нейролеммоцитами. Эти клетки подвижны и могут даже перекочевывать с одного отростка нервной клетки на другой. Они, распластываясь на поверхности отростка нервной клетки, постепенно скользят по нему. Было установлено, что леммоцит, уплощаясь, постепенно охватывает отросток нервной клетки и смыкается. Место контакта краев клетки называется мезаксоном, т.е. мезаксон представляет собой место соединения двух цитолемм. Иногда шванновская клетка охватывает несколько отростков нервных клеток, в результате образуются нервные волокна кабельного типа. Таким образом, безмиелиновые нервные волокна состоят из осевого цилиндра и глиальной или шванновской непрерывной оболочки. При световой микроскопии безмиелиновые нервные волокна имеют вид тонких тяжей и многочисленных просвечивающихся ядер. Границы шванновских клеток очень тонкие, поэтому они не видны. Рост аксонов происходит поградиенту концентрации специфических химических факторов, вырабатываемых в мишенях (например, фактор роста нервов; ацетилхолин определяет направление роста аксона). Кроме того, возможно, что в пространстве роста аксона распределены молекулярные метки, которые считываются друг за другом растущим отростком, в результате чего он растет в нужном направлении.
Скорость проведения нервного импульса по безмиелиновым нервным волокнам до 5 метров в секунду.
Миелиновые нервные волокна встречаются преимущественно в центральной нервной системе. Первоначально миелиновые волокна образуются точно также как и безмиелиновых волокон. Однако после образования мезаксона развитие безмиелиновых нервных волокон завершается. При образовании миелинового нервного волокна после образования мезаксона клетка начинает вращаться вокруг отростка нервной клетки, в результате чего мезаксон наматывается на отросток, а цитоплазма шванновской клетки оттесняется на периферию. За счет намоток мезаксона образуется дополнительная оболочка нервного волокна, которая получила название миелиновая оболочка. Слои поверхностной мембраны шванновской клетки содержат белки и липоиды, поэтому при многократном наслаивании мезаксона образуется темная миелиновая оболочка, состоящая из холестерина, нейтральных жиров и фосфатидов. Таким образом, миелиновое нервное волокно состоит из осевого цилиндра, окруженного миелиновой и шванновской оболочками. При световой микроскопии на срезах, обработанных осмием, видно, что миелиновое нервное волокно состоит из темной прерывистой миелиновой оболочки и очень тонкой непрерывной шванновской оболочки. Участки, где миелиновая оболочка прерывается, нервное волокно истончается. Эти участки получили название перехватов Ранвье. Таким образом на месте перехвата Ранвье осевой цилиндр покрыт только неврилеммой (шванновской оболочкой). Расстояние между двумя перехватами Ранвье соответствует границам одной шванновской клетки, содержащей одно или два ядра. В области перехвата Ранвье шванновские клетки дают многочисленные пальцевидные выросты, которые беспорядочно переплетаются. Плазматическая мембрана осевого цилиндра в области перехвата Ранвье отличается высокой концентрацией ионных каналов, в особенности натриевых, что обеспечивает генерацию и проведение потенциала действия по длине осевого цилиндра. Миелиновая оболочка неоднородна: в ее толще обнаруживаются насечки Шмидта-Лантермана, которые видны в виде светлых полосок, пересекающих миелиновую оболочку в косом направлении. При электронной микроскопии насечки видны в виде участков, где мембраны имеют неправильный ход или складки. Значение этого явления не установлено. Скорость проведения нервного импульса по миелиновым волокнам достигает 120 метров в секунду, благодаря скачкообразному проведению импульса. Миелиновая оболочка изолирует аксон от индуцирующего влияния со стороны соседних нервных волокон.
Развитие миелиновых волокон в разных участках происходит в разное время. Было показано, что филогенетически более старые системы проводников одеваются миелином раньше. Процесс миелинизации нервных волокон не заканчивается к рождению и продолжается первые годы жизни ребенка. Так, процесс миелинизации черепно-мозговых нервных волокон заканчивается только к 1-1,5 годам, а миелинизация спинальных нервов может растягиваться до 5 лет. Развитие миелиновых оболочек особенно усиливается у ребенка с 8 месяцев жизни в период начала ходьбы. При этом, миелинизация двигательных нервных волокон идет быстрее, чем чувствительных.
Нервные волокна на периферии редко идут одиночно, изолированно. Чаще они лежат пучками, образуя нервы.
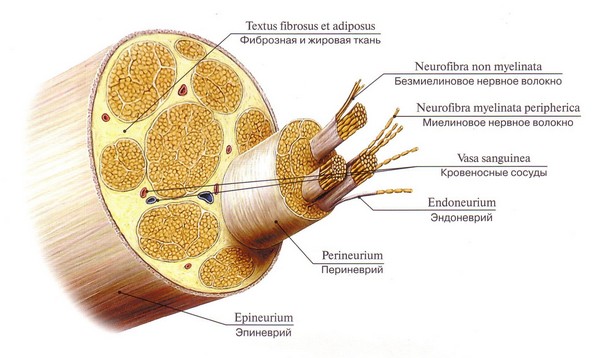
Периферический нерв состоит как из миелиновых, так и из безмиелиновых нервных волокон. При этом, те или иные нервные волокна могут преобладать в периферическом нерве. В составе периферического нерва каждое нервное волокно окружено очень тонкой прослойкой нежной соединительной ткани, содержащей кровеносные сосуды. Это эндоневрий. Кровеносные сосуды эндоневрия разветвляются на многочисленные капилляры, которые обеспечивают питание нервных волокон. Отдельные пучки нервных волокон в составе периферического нерва разграничены более выраженными прослойками рыхлой соединительной ткани, которые получили название периневрий. Периневрий с внутренней поверхности выстлан несколькими слоями (от 3 до 10) уплощенных эпителиальных клеток, способных к фагоцитозу. Установлено, что они могут фагоцитировать лепрозные бактерии. По мере истончения нервов число слоев эпителиальных клеток уменьшается, вплоть до одного слоя. В соединительной ткани периневрия содержатся фибробласты, тучные клетки. На обеих поверхностях каждого эпителиального слоя располагается базальная мембрана. Последний эпителиальный слой исчезает вместе со шванновскими клетками при образовании терминалей. Шванновские и эпителиальные клетки периневрия имеют общую ультраструктурную характеристику, но обладают разными антигенными свойствами. Периневрий выполняет барьерную функцию, так как обладает избирательной проницаемостью для различных красителей, коллоидов, протеинов, пероксидазы хрена, электролитов, то есть образует гемато-нейральный барьер, который функционально и структурно соответствует гемато-энцефалическому барьеру центральной нервной системы. Периневрий принимает активное участие в процессах регенерации нервных волокон. Так, установлено, что при повреждении периневрия регенерация нервного волокна не происходит.
С поверхности периферический нерв покрыт эпиневрием, состоящим из коллагеновых и даже эластических волокон. Здесь проходят кровеносные сосуды и залегают отдельные скопления жировых клеток.
Регенерация нервных волокон. Развивающиеся при травме деструктивные и дегенеративные субклеточные процессы, стимулируют одновременно и процессы восстановления.
При повреждении мякотных нервных волокон развивается валлеровская дегенерация, которая наступает уже через 3-7 часов после травмы. Она характеризуется появлением неровности контуров нервного волокна и распада и расслоение миелина на отдельные фрагменты и ее вакуолизация. Миелин распадается до нейтрального жира. Распад миелиновой оболочки происходит до нейтральных жиров. Распад миелина идет параллельно с разрушением (некрозом) осевых цилиндров. Продукты их распада в течение нескольких месяцев резорбируются шванновскими клетками и макрофагами эндоневрия и периневрия (поглощаются, перевариваются и рассасываются). В перикарионе травмированных нейронов наблюдается уменьшение числа канальцев гранулярной эндоплазматической сети (тигролиз). В последующем на месте дегенерировавших участков миелиновых и безмиелиновых нервных волокон остаются лишь тяжи шванновских клеток (бюнгнеровские ленты), которые интенсивно пролиферируют прорастают на встречу друг другу из обоих концов нерва. Одновременно идет разрастание соединительной ткани и кровеносных сосудов. Уже через 3 часа после травмы на концах поврежденных участков (центрального и периферического) образуются утолщения- натеки аксоплазмы, получившие название колбы роста (концевые колбы). Благодаря способности тела нервной клетки продуцировать аксоплазму, от колб роста начинают расти многочисленные безмиелиновые коллатерали, на концах которых формируются колбы, натеки, спирали, намотки, шары. Образовавшиеся коллатерали постепенно продвигаются к перерезанному концу аксона в области травмированной зоны. При этом, часть коллатералей дегенерирует, а остальные продолжают свой рост к периферическому концу нерва. Установлено, что успешная регенерация происходит в том случае, если в периферический конец нерва прорастает достаточное количество аксонов для восстановления нервных связей с рабочими органами. Одновременно происходит интенсивная пролиферация шванновских клеток, что приводит в конечном итоге к образованию мощных скоплений глиальных клеток. Коллатерали прорастают пласт шванновских клеток и покрываются ими, приобретая при этом глиальную оболочку.
Скорость регенерации аксонов периферических нервных волокон происходит у человека 0,1-1,5 мм в сутки (редко до 5 мм в сутки). У детей регенерация идет значительно быстрее. Регенерирующие безмиелиновые нервные волокна через 20-30 дней после травмы покрываются миелиновой оболочкой. Однако обычной толщины она достигает только через 6-8 месяцев после травмы. Степень реиннервации нервного ствола определяется по количеству прорастающих в него нервных волокон. Рост аксонов происходит по градиенту концентрации специфических химических факторов, вырабатываемых в мишенях, например, фактор роста нервов. Большое значение для восстановления аксонов имеют сохранившиеся шванновские клетки, которые маркируют направление роста отростка. Растущий отросток двигается по поверхности этих клеток между плазмолеммой и базальной мембраной. Выделяемые шванновскими клетками нейротрофические факторы, в том числе фактор нервов, поглощаются аксоном и транспортируются в перикарион, где стимулируют синтез белка. Предполагают, что в пространстве роста аксона распределены молекулярные метки. Растущий отросток считывает одну за другой метки и растет в нужном направлении. Если аксон не находит путь роста по шванновским клеткам, то наблюдается хаотичное разрастание его разветвлений.
Основным препятствием для регенерации аксонов поврежденного нерва служит грубый соединительнотканный рубец, образующийся в зоне травмы. В связи с этим, чтобы избежать различного рода осложнений, возникающих в месте травмы, нарушения кровообращения, улучшить регенерацию используются оптимальные методы обработки раны, современные виды шовного материала для соединения концов нерва. Так, предложен полимерный клей, который образовывает вокруг эпиневрия своеобразную муфту, что обусловливает развитие рыхлого соединительнотканного рубца, который в меньшей степени препятствует регенерации. Кроме того, установлено, что твердая мозговая оболочка обладает очень низкой антигенной активностью и быстро рассасывается в тканях, вызывая минимальные воспалительные изменения. В связи с этим, предложено использование твердой мозговой оболочки для изоляции места травмы периферических нервов от окружающих тканей и нитей из нее в качестве шовного материала, что существенно улучшило лечение больных. Кроме того, для ускорения регенерации используются и другие методы. Например, концы поврежденного нерва помещают в трубки, в которые заливают аутогенную сыворотку, благодаря чему снижается инвазия фибробластов. «Метод естественного резерва длины» позволяет без вреда вытянуть поврежденный нерв, так как он располагается зигзагообразно. Применяется аутопластика, то есть пересадка в область травмы отрезка другого нерва. Иногда используют культуру шванновских клеток, которую помещают в область травмы.
Отростки нервных клеток, аксоны или дендриты, заканчиваются либо в тканях, где образуют нервные окончания, либо контактируют с другими клетками, образуя синапсы.
Синапсы представляют собой сложные структуры, формирующиеся в области контакта двух клеток, специализирующиеся на одностороннем проведении нервного импульса.
Понятие синапса было введено на основании физиологических наблюдений Шеррингтоном в 1897 году. Окончательное подтверждение их наличия было осуществлено только в середине 20 столетия с помощью электронного микроскопа. Тем самым была завершена многолетняя дискуссия между сторонниками «нейронной теории» строения нервной системы, согласно которой, нервная клетка считалась основной структурной и функциональной единицей, и сторонниками теории «контуитета», которые провозглашали постулат о непрерывном соединении нейрофибрилл между отростками клеток в единую сеть. Синапсы обладают высокой пластичностью. В головном мозге человека насчитывается 10 химических синапсов.
По характеру контакта различают несколько видов синапсов: аксо-соматические, аксо-дендритические, аксо-аксональные, дендро-дендритические, дендро-соматические (последние три вида синапсов являются тормозными).
По локализации различают центральные синапсы, расположенные в центральной нервной системе, и периферические, лежащие в периферической нервной системе, в том числе в вегетативных ганглиях.
По развитию в онтогенезе различают статические синапсы, расположенные в рефлекторной дуге безусловных рефлексов, и динамические, характерные для рефлекторных дуг условных рефлексов.
По конечному эффекту различают возбуждающие синапсы и тормозные синапсы.
По механизму передачи нервного импульса различают электрические синапсы, химические синапсы и смешанные синапсы. Электрический синапс отличается прежде всего своей симметричностью и тесными контактами обеих мембран. Суженная синаптическая щель в месте электрического контакта перекрыта тонкими канальцами, через которые осуществляется быстрое продвижение ионов между нервными клетками. Таким образом, электрический синапс представляет собой щелевидный контакт между двумя клетками с ионными каналами. Аналогом электрического синапса у человека являются щелевидные контакты в сердечной мышечной ткани. Все синапсы у человека являются практически химическими, так как используют для передачи нервного импульса с одной клетки на другую химическое соединение: нейромедиатор или нейротрансмиттер.
По характеру нейротрансмиттера различают синапсы: холинергические, использующие в качестве нейромедиатора ацетилхолин, адренергические (норадреналин), дофаминергические (дофамин), ГАМК-ергические (ГАМК), пептидергические (пептиды), пуринергические (АТФ). Например, при шизофрении увеличивается число синапсов, использующих для передачи импульса дофамин. В качестве нейротрансмиттеров могут использоваться глутамат, гистамин, серотонин, глицин. В настоящее время принято считать, что каждый нейрон продуцирует более одного нейротрансмиттера.
В области контакта плазмолемма аксона утолщается и называется пресинаптической мембраной. В аксоплазме содержатся многочисленные митохондрии и синаптические пузырьки, содержащие нейротрансмиттер- ацетилхолин (или другой медиатор). Плазмолемма другой клетки в области контакта также утолщается и называется постсинаптической мембраной. Узкое щелевидное пространство между этими мембранами представляет собой синаптическую щель. В пресинаптической мембране имеются многочисленные кальциевые каналы, которые открываются при прохождении волны деполяризации. Постсинаптическая мембрана содержит холинорецепторы, проявляюшие высокую чувствительность к ацетилхолину. При деполяризации пресинаптической мембраны кальциевые каналы открываются и ионы кальция выходят, запуская выход в синаптическую щель ацетилхолина. В каждом синаптическом пузырьке содержится несколько тысяч молекул нейромедиатора, что составляет квант. Синаптические пузырьки могут сливаться с постсинаптической мембраной только при повышении концентрации ионов кальция. В настоящее время синтезирован целый ряд лекарственных препаратов, блокирующих кальциевые каналы, которые широко используются в кардиологии при лечении некоторых видов аритмий. Квант ацетилхолина достигает поверхности постсинаптической мембраны и взаимодействует с холинорецепторами. В результате взаимодействия ацетилхолина с холинорецептором рецепторный белок меняет свою конфигурацию, что приводит к повышению проницаемости постсинаптической мембраны для ионов. Это обусловливает перераспределение ионов калия и натрия по обе стороны мембраны и возникновение волны деполяризации.
Устранение ацетилхолина в дальнейшем происходит за счет ацетилхолинэстеразы, локализованной в синапсе. Ряд химических соединений, в том числе фосфорорганические соединения, токсины бледной поганки ингибируют холинэстеразу, что приводит к высокой концентрации ацетилхолина в синаптической щели, поэтому в этих случаях вводят антидот- атропин, который блокирует холинорецепторы.
Нервные волокна в тканях заканчиваются нервными окончаниями, которые представляют собой сложные структуры на концах дендритов и аксонов в тканях. Все нервные окончания подразделяются на два вида: чувствительные и двигательные.
Чувствительные нервные окончания или рецепторы образованы дендритами нервных клеток. По локализации различают экстерорецепторы, воспринимающие информацию от покровных тканей (например, рецепторы кожи, слизистых оболочек) и интерорецепторы, воспринимающие информацию от внутренних органов (например, рецепторы сосудов). По характеру воспринимающего раздражения различают терморецепторы, хеморецепторы, механорецепторы, барорецепторы, нацирецепторы и др.
По строению рецепторы подразделяются на свободные и несвободные (классификация Лаврентьева). Свободные рецепторы представляют собой структуры в образовании которых участвует только осевой цилиндр, то есть они свободны от клеток глии (если быть точным, то шванновские клетки присутствуют в очень небольшом количестве). При этом ветвления осевого цилиндра лежат свободно среди клеток эпителия. Свободные рецепторы, как правило, воспринимают болевые ощущения.
Несвободные рецепторы образуются ветвлением осевого цилиндра, которые сопровождаются клетками глии, то есть они не свободны от клеток глии. Несвободные рецепторы подразделяются на инкапсулированные и рецепторы с дополнительными структурами.
Инкапсулированные нервные окончания характеризуются наличием сложных оболочек. К инкапсулированным нервным окончаниям относятся пластинчатые тельца (тельца Фатер-Пачини) и осязательные тельца Мейснера. Тельца Фатер-Пачини характерны для соединительно ткани, по характеру воспринимаемого раздражения являются барорецепторами. При образовании этого нервного окончания миелиновое нервное волокно теряет миелиновую оболочку, оставшийся осевой цилиндр разветвляется, его ветвления сопровождаются небольшим количеством глиальных клеток. С поверхности тельце Фатер-Пачини окружено соединительнотканной касулой, состоящей из многочисленных пластинок, наслаивающихся друг на друга. Каждая пластинка состоит из тонких коллагеновых волокон, склееных аморфным веществом, и фибробластов, лежащих между ними.
К инкапсулированным нервным окончаниям относятся также осязательные тельца Мейснера, находящиеся в составе сосочков кожи. Миелиновое нервное волокно, подойдя к сосочку кожи, теряет миелиновую оболочку и обильно разветвляется между многочисленными клетками олигодендроглии. С поверхности тельце покрыто тонкой соединительнотканной капсулой, состоящей в основном из тонких коллагеновых волокон.
К рецепторам с дополнительными структурами относятся диски Меркеля, которые находятся в кожном эпителии. Они представлены клетками Меркеля и контактирующими с ними дендритами нервных клеток. Клетка Меркеля представляет собой видоизмененную эпителиальную клетку(светлая цитоплазма, уплощенное ядро, многочисленные осмиофильные гранулы), лежащую в составе эпителия. Вокруг клетки Меркеля располагается спирально закрученные дендритные веточки. Диски Меркеля обеспечивают высокую тактильную чувствительность.
В скелетной мышечной ткани чувствительные нервные окончания представлены нервно-мышечными веретенами, регистрирующие изменения длины мышечных волокон и скорость их изменений. Веретено состоит из нескольких (до 10-12) тонких и коротких поперечнополосатых мышечных волокон, окруженных тонкой растяжимой капсулой. Это интрафузальные волокна. Волокна, лежащие за пределами капсулы называются экстрафузальными. Актиновые и миозиновые миофибриллы содержатся только на концах интрафузальных волокон, поэтому сокращаться могут только концы интрафузальных мышечных волокон. При этом центральная часть интрафузальных мышечных волокон является несокращающейся. Она является рецепторной. Различают два вида интрафузальных мышечных волокон: волокна с ядерной цепочкой и с ядерной сумкой. Волокон с ядерной сумкой в каждом веретене от 1 до 3. Центральная их часть расширена и содержит много ядер. Волокон с ядерной цепочкой может быть в веретене от 3 до 7. Эти волокна в два раза тоньше и более короткие, а ядра в них расположены цепочкой по всей рецепторной части. К интрафузальным мышечным волокнам подходят афферентные волокна двух типов. Одни из них образуют окончания в виде спирали, оплетающие интрафузальные волокна. Другие образуют гроздьевидные окончания, которые лежат по обе стороны от спиральных окончаний. Когда мышца расслабляется или сокращается происходит изменение длины интрафузальных волокон, что регистрируется рецепторами. Спиральные окончания регистрируют изменение длины мышечного волокна и скорость этого изменения, а гроздьевидные окончания регистрируют только изменение длины. Эфферентная иннервация представлена аксомышечным синапсом на концах мышечного волокна. Вызывая сокращение концевых участков интрафузального мышечного волокна, они обусловливают растяжение его центральной рецепторной части.
Двигательные нервные окончания образованы концевыми отделами аксонов нервных клеток спинного мозга. При световой микроскопии двигательные нервные окончания (эффекторы) имеют вид кустиков или птичьих лапок с пуговчатыми утолщениями на концах. Важно, что двигательные нервные окончания кроме передачи нервного импульса они оказывают трофическое действие, регулируя метаболизм клеток и тканей. При электронной микроскопии эффекторы построены по типу синапса.
Двигательные окончания в скелетных мышцах называются моторными бляшками. Моторная бляшка состоит из из концевого ветвления аксона и подошвы. Миелиновое нервное волокно, подходя к мышечному волокну, теряет миелиновую оболочку и прогибает сарколемму в виде многочисленных пальцевидных выростов. В сарколемме, которая образует впячивания, возникают еще более мелкие углубления. Неврилемма аксона срастается с сарколеммой и возникает конусообразное пространство, заполненное цитоплазмой леммоцитов, здесь же лежат и ядра. В этом пространстве ветвится осевой цилиндр. Пресинаптическая оболочка представлена в двигательной бляшке аксолеммой. Постсинаптической мембраной является сарколемма мышечного волокна. Между этими мембранами формируется щелевидное пространство- синаптическая щель. В нейроплазме аксона концентрируется много митохондрий и мелкие синаптические пузырьки. В саркоплазме мышечного волокна в области бляшки также наблюдается скопление ядер.
Особенности нервных волокон и нервных окончаний в детском организме.
Нервные волокна. В период новорожденности нервные волокна более короткие и более тонкие, чем у взрослого человека. Возрастными особенностями строения периферических нервных волокон является этапный характер их миелинизации. Миелинизация нервных волокон начинается еще во внутриутробном периоде. Первыми миелинизируются волокна филогенетически более древних жизненно важных органов и систем. Однако к рождению ребенка миелинизация не заканчивается. К 9 годам миелинизация нервных волокон в периферических нервах близка к завершению. Миелинизация черепномозговых нервов заканчивается к 1,5 годам, а спинномозговых только к 5 годам. Миелинизация двигательных нервных волокон идет быстрее, чем чувствительных. Миелинизация волокна происходит в центробежном направлении, то есть от клетки к терминалям. Расстояние между перехватами Ранвье у ребенка значительно меньше, чем у взрослого. С возрастом толщина миелиновой оболочки увеличивается. До 3-х лет у ребенка прослойки соединительной ткани более выражены и богаты клеточными элементами.
Нервное волокно – это удлиненный отросток нейронов, покрытый леммоцитами и миелиновой или безмиелиновой оболочкой. Основной его функцией является проводимость . В периферической и центральной нервной системе преобладают мякотные (миелиновые) нервные волокна, которые иннервируют скелетную мускулатуру, безмякотные находятся в симпатическом отделе вегетативной системы и распространяются на внутренние органы. Волокна, не имеющие оболочки, называются голыми осевыми цилиндрами.
Нервное волокно имеет в основе отросток нейрона, который образует своеобразную ось. Снаружи он окружен миелиновой оболочкой с биомолекулярной липидной основой, состоящей из большого количества витков мезаксона, который по спирали накручивается на нейроновую ось. Таким образом, происходит миелинизация нервных волокон.
Миелиновые нервные волокна периферической системы сверху дополнительно покрыты вспомогательными Шванновскими клетками, поддерживающими аксон и питающими тело нейрона. Поверхность мякотной мембраны имеет интервалы – перехваты Ранвье, в этих местах осевой цилиндр прикрепляется к наружной Шванновской мембране.
Миелиновый слой не обладает электропроводящими свойствами, их имеют перехваты. Возбуждение происходит в ближайшем к месту воздействия внешнего раздражителя интервале Ранвье. Импульс передается скачкообразно, от одного перехвата к другому, это обеспечивает высокую скорость распространения импульса.
Миелиновые нервные волокна регулируют обмен веществ в мышечной ткани, обладают высоким сопротивлением по отношению к биоэлектрическому току.
Промежутки Ранвье генерируют и усиливают импульсы. У волокон центральной нервной системы нет Шванновской мембраны, эту функцию выполняют олигодендроглии.
Безмякотные ткани имеют несколько осевых цилиндров, у них нет миелинового слоя и перехватов, сверху покрыты Шванновскими клетками, между ними и цилиндрами образуются щелевидные пространства. Волокна имеют слабую изоляцию, допускают распространение импульса из одного отростка нейрона в другой, на всем протяжении контактируют с окружающей средой, скорость проведения импульсов гораздо ниже, чем у мякотных волокон, при этом организму требуется большее количество энергии.
Из мякотных и безмякотных отростков нейронов формируются крупные нервные стволы, которые, в свою очередь, разветвляются на более мелкие пучки и заканчиваются нервными окончаниями (рецепторные, двигательные, синапсы).
Нервные окончания – это конец миелиновых и безмиелиновых нервных волокон, который формирует межнейронные контакты, рецепторные и двигательные окончания.
Принципы классификации

Разные типы нервных волокон имеют неодинаковую скорость проведения импульсов возбуждения, это зависит от их диаметра, длительности потенциала действия и степени миелинизации. Существует прямо пропорциональная зависимость между скоростью и диаметром волокна.
Структурно-функциональный метод классификации нервных волокон Эрлангера-Гассера по :
- Миелиновое нервное волокно группы А: α, β, Υи δ. Самый большой диаметр и толстую оболочку имеют ткани α – 20 мк, они обладают хорошей скорость проводимости импульсов – 120 м/сек. Эти ткани иннервируют источник возбуждения из столба спинного мозга к скелетным рецепторам мышц, сухожильям, отвечают за тактильные ощущения.
Остальные типы волокон имеют меньший диаметр (12 мк), скорость проведения импульса. Эти ткани передают сигналы от внутренних органов, источников боли в ЦНС.
- Миелиновые волокна группы В относятся к . Общая скорость проведения импульса составляет 14 м/сек, потенциал действия в 2 раза больше, чем у волокон группы А. Миелиновая оболочка слабо выражена.
- Безмиелиновые волокна группы С имеют очень маленький диаметр (0,5 мк) и скорость возбуждения (6 м/сек). Эти ткани иннервируют . К данной группе также относятся волокна, которые проводят импульсы от центров боли, холода, тепла и давления.
Отростки нейронов делят на афферентные и эфферентные. Первый тип обеспечивает передачу импульсов от рецепторов тканей в центральную нервную систему. Второй тип передает возбуждение от ЦНС к рецепторам тканей.
Функциональная классификация нервных волокон афферентного типа по Ллойду-Ханту:
Демиенилизация
Процесс демиелинизации нервных волокон – это патологическое повреждение миелиновой оболочки, которое вызывает нарушение функционирования тканей. Вызывают патологию воспалительные процессы, метаболические нарушения, нейроинфекция, интоксикация или ишемия тканей. Миелин замещается фиброзными бляшками, в результате нарушается проведение импульсов.
Первый тип демиелинизации – это миелинопатия, вызванная аутоиммунными реакциями организма, болезнью Канавана, амиотрофией Шарко-Мари-Тута.
Второй тип – это миелинокластия. Патология характеризуется наследственной предрасположенностью к разрушению миелиновой оболочки (болезнь Бинсвангера).
Демиелинизирующие заболевания

Заболевания, приводящие к разрушению миелиновой оболочки, чаще всего имеют аутоиммунную природу, другой причиной может быть лечение нейролептиками или наследственная предрасположенность. Разрушение липидного слоя вызывает снижение скорости проведения импульсов раздражения.
Заболевания разделяют на те, которые затрагивают центральную нервную систему и патологии, повреждающие периферическую сеть. Болезни, которые влияют на работу ЦНС:
- Миелопатия спинного мозга возникает в результате сдавливания миелиновых волокон межпозвоночными грыжами, опухолями, костными осколками, после . У больных снижается чувствительность, мышечная сила в области поражения, возникают парезы рук или ног, нарушается работа кишечника, мочевыводящей системы, развивается атрофия мышц нижних конечностей.
- Лейкодистрофия головного мозга вызывает поражение белого вещества. У пациентов нарушена координация движений, они не могут держать равновесие. Развивается мышечная слабость, появляются непроизвольные судороги, . Постепенно ухудшается память, интеллектуальные способности, зрение и слух. На поздних стадиях возникает слепота, глухота, полный паралич, трудности во время проглатывания пищи.
- головного мозга чаще всего поражает мужчин старше 60 лет. Основными причинами является артериальная гипертензия и наследственная предрасположенность. У пациентов ухудшается память и внимание, появляется заторможенность, трудности с речью. Замедляется походка, нарушается координация движений, появляется недержание мочи, больному тяжело глотать пищу.
- Синдром осмотической демиелинизации характеризуется распадом миелиновых оболочек в тканях головного мозга. У больных отмечается расстройство речевого аппарата, постоянное чувство сонливости, депрессии или повышенная возбудимость, мутизм, парез всех конечностей. На ранних стадиях заболевания процесс демиелинизации обратим.
- Рассеянный склероз проявляется онемением одной или двух конечностей, частичная или полная потеря зрения, боль при движении глаз, головокружение, быстрая утомляемость, тремор конечностей, нарушение координации движений, покалывание в различных частях тела.
- Болезнь Девика – это воспалительный аутоиммунный недуг, который поражает зрительный нерв и ствол спинного мозга. К симптомам относится различная степень нарушения зрения, вплоть до слепоты, парапарезы, тетрапарезы, нарушение функционирования органов малого таза.
Симптомы заболеваний зависят от области поражения миелиновых волокон. Выявить процесс демиелинизации можно с помощью компьютерной томографии, магниторезонансной терапии. Признаки обнаруживаются на электромиографии.
МИЕЛИНИЗАЦИЯ , процесс обложения миелином нервного волокна в период развития организма (см. отдельную таблицу, рисунки 1-3). М. начинается у.зародыша на 5-м месяце внутриутробной жизни; отделы мозга миелинизируются не одновременно, а в известном закономерном порядке. Системы волокон, имеющие одинаковую по сложности функцию, миелинизируются одновременно; чем сложнее функция данной системы, тем волокна ее позднее обкладываются миелином; обложение миелином служит признаком того, что волокно стало деятельным. При рождении ребенка М. далеко еще не закончена: в то время как одни части мозга уже вполне миелинизированы и готовы к функции, другие еще не закончили своего развития й не могут служить ни для физ. ни для псих, отправлений.У новорожденного ребенка спинной мозг очень богат миелино-выми волокнами; необложенные миелином волокна находятся только во внутренних его частях и в области пирамидного пучка. Волокна мозгового ствола и мозжечка в значительном количестве покрыты миелиновой оболочкой. Из подкорковых узлов волокна globi pallidi уже миелинизированы, тогда как волокна nucl. caudati и putamen покрываются миелином только к 5-6 месяцам внеутробной жизни. Полушария большого мозга во многих своих частях лишены миелина и на разрезе имеют сероватый цвет: у нормального новорожденного ребенка миелином снабжены центростремительные (чувствующие) волокна, часть пирамидных путей, часть обонятельных, слуховых и зрительных путей и центров и отдельные участки в corona radiata; большая же часть теменных, лобных, височных и затылочных долей, равно как и комиссур полушария, еще лишены миелина. Ассоциационные системы, назначенные для высших, псих, функций, обкладываются миелином позднее других систем, благодаря чему корковые зоны проекционных центров и волокон остаются изолированными, не связанными между собой; в этот период все ощущения, получаемые ребенком извне, остаются изолированными, все движения его рефлекторны и появляются только вследствие внешних или внутренних раздражений. Постепенно развитие миелиновых оболочек происходит во всех отделах мозга, благодаря чему устанавливается связь между различными центрами и в связи с этим развивается интелект ребенка: он начинает узнавать предметы и понимать их значение. Миелинизация главных систем полушария заканчивается на восьмом месяце внеутробной жизни, и с этого момента она продолжается только в отдельных волокнах в течение еще многих лет (наружные слои мозговой коры по нек-рым данным миелинизируются окончательно лишь к 45 годам жизни и м. б. даже позднее). В зависимости от времени появления миелина в полушариях мозга Флексиг (Flechsig) делит их на разные области: те части, где волокна покры- ваются миелином рано, он называет ранними областями (Primordialgebiete), те же, в к-рых миелин появляется позднее, - поздними (Spatgebiete). На основании этих исследований Флексиг различает в коре головного мозга двоякого рода центры: одни соединены проекционными волокнами с нижележащими образованиями, это - проекционные центры;"другие, не имеющие связи с нижележащими отделами мозга, но связанные ассоциационными волокнами с проекционными центрами коры, являются а с с о-циационными центрами (см. Голов-ной мозг, т. VII, ст. 533-534). При изучении головнбго мозга миелинизацией пользуются как методом-метод миелогенетический или метод Флексига. Лит.: Бехтерев В., Проводящие пути головного и спинного мозга, СПБ, 1896; Flechsig F., Anatomie des menschlichen G-ehirns und Ruckenmarks auf myelogenetischer Grundlage, Lpz., 1920 (лит.); PfeiferR., Myelogenetiscn-anatomische Untersu-chungen uber den zentralen Abschnitt der Sehleitung (Monographien aus dem G-esamtgebiete der Neurologie und Psvchiatrie, hrsg. v. O. Foerster u. K. Wilmanns, B. XLIII, В., 1925).E. Кононова.
Обеспечивается олигодендроцитами. Каждый олигодендроглиоцит образует несколько «ножек», каждая из которых оборачивает часть какого-либо аксона. В результате один олигодендроцит связан с несколькими нейронами. Перехваты Ранвье здесь шире, чем на периферии. Согласно исследованию 2011 г. мощную миелиновую изоляцию в мозге получают наиболее активные аксоны, что позволяет им далее работать ещё эффективнее. Важную роль в этом процессе играет сигнализатор глутамат.
в миелинизированные волокна в НС проводят импульс быстрее, чем немиелинизоровнные
Миелиновая оболочка - это не клеточная мембрана. Оболочку образуют шванновские клетки, типа рулета, они создают области с высоким сопротивлением, и ослабляют ток утечки из аксона. Получается, что потенциал как бы перескакивает от перехваток перехвату, от этого и скорость проведения импульса становится выше.
8. Си́напс (греч. σύναψις, от συνάπτειν - обнимать, обхватывать, пожимать руку) - место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками , причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться.
Типичный синапс - аксо-дендритический химический. Такой синапс состоит из двух частей: пресинаптической
, образованной булавовидным расширением окончаниема ксона передающей клетки и постсинаптической
, представленной контактирующим участком цитолеммы воспринимающей клетки (в данном случае - участком дендрита). Синапс представляет собой пространство, разделяющее мембраны контактирующих клеток, к которым подходят нервные окончания. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую. 
9. Химический синапс - особый тип межклеточного контакта между нейроном и клеткой-мишенью. Состоит из трёх основных частей: нервного окончания с пресинаптической мембраной , постсинаптической мембраны клетки-мишени и синаптической щели между ними.
электрические - клетки соединяются высокопроницаемыми контактами с помощью особых коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе - 3,5 нм (обычное межклеточное - 20 нм).Так как сопротивление внеклеточной жидкости мало(в данном случае), импульсы проходят не задерживаясь через синапс. Электрические синапсы обычно бывают возбуждающими.
При деполяризации пресинаптической терминали открываются потенциал-чувствительные кальциевые каналы, ионы кальция входят в пресинаптическую терминаль и запускают механизм слияния синаптических пузырьков с мембраной. В результате медиатор выходит в синаптическую щель и присоединяется к белкам-рецепторам постсинаптической мембраны, которые делятся на метаботропные и ионотропные. Первые связаны с G-белком и запускают каскад реакций внутриклеточной передачи сигнала. Вторые связаны с ионными каналами, которые открываются при связывании с ними нейромедиатора, что приводит к изменению мембранного потенциала. Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели - ацетилхолинэстераза. Одновременно часть медиатора может перемещаться с помощью белков-переносчиков через постсинаптическую мембрану (прямой захват) и в обратном направлении через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
10. Нервно-мышечный синапс (мионевральный синапс) - эффекторное нервное окончание на скелетном мышечном волокне.
Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с плазматической мембраной мышечного волокна, образующийся из выпячиваний аксона и цитолеммы мышечного волокна, создавая глубокие «карманы». Синаптическая мембрана аксона и постсинаптическая мембрана мышечного волокна разделены синаптической щелью. В этой области мышечное волокно не имеет поперечной исчерченности, характерно скопление митохондрий и ядер. Терминали аксонов содержат большое количество митохондрий и синаптических пузырьков с медиатором (ацетилхолином).
1. Пресинаптическое окончание 
2. Сарколемма
3. Синаптический пузырек
4. Никотиновый ацетилхолиновый рецептор
5. Митохондрия
11. Нейромедиа́торы (нейротрансмиттеры , посредники ) - биологически активные химические вещества, посредством которых осуществляется передача электрического импульса с нервной клетки через синаптическое пространство между нейронами . Нервный импульс, поступающий в пресинаптическое окончание, вызывает освобождение в синаптическую щель медиатора. Молекулы медиаторов реагируют со специфическими рецепторными белками клеточной мембраны, инициируя цепь биохимических реакций, вызывающих изменение трансмембранного тока ионов, что приводит к деполяризации мембраны и возникновению потенциала действия.
Нейромедиаторы являются, как и гормоны, первичными мессенджерами, но их высвобождение и механизм действия в химических синапсах сильно отличается от такового гормонов. В пресинаптической клетке везикулы, содержащие нейромедиатор, высвобождают его локально в очень маленький объём синаптической щели. Высвобожденный нейромедиатор затем диффундирует через щель и связывается с рецепторами на постсинаптической мембране. Диффузия является медленным процессом, но пересечение такой короткой дистанции, которая разделяет пре- и постсинаптические мембраны (0,1 мкм или меньше), происходит достаточно быстро и позволяет осуществлять быструю передачу сигнала между нейронами или между нейроном и мышцей.
Недостаток какого-либо из нейромедиаторов может вызывать разнообразные нарушения, например, различные виды депрессии. Также считается , что формирование зависимости от наркотиков и табака связано с тем, что при употреблении этих веществ задействуются механизмы производства нейромедиатора серотонина, а также других нейромедиаторов, блокирующие (вытесняющие) аналогичные естественные механизмы.
Классификация нейромедиаторов:
Традиционно нейромедиаторы относят к 3 группам: аминокислоты, пептиды, моноамины (в том числе катехоламины)
Аминокислоты :
§ Глутаминовая кислота (глутамат)
Катехоламины :
§ Адреналин
§ Норадреналин
§ Дофамин
Другие моноамины :
§ Серотонин
§ Гистамин
А также :
§ Ацетилхолин
§ Анандамид
§ Аспартат
§ Вазоактивный интестинальный пептид
§ Окситоцин
§ Триптамин
12. Нейроглия, или просто глия - сложный комплекс вспомогательных клеток нервной ткани, общный функциями и, частично, происхождением (исключение - микроглия).Глиальные клетки составляют специфическое микроокружение для нейронов, обеспечивая условия для генерации и передачи нервных импульсов, обеспечивают тканевый гомеостаз и нормальное функц-е клетки, а также осуществляя часть метаболических процессов самого нейрона. Основные функции Нейроглии:
Создание между кровью и нейронами гемато-энцефалического барьера, необходимого как для защиты нейронов, так и главным образом для регуляции поступления веществ в ЦНС и их выведения в кровь;
Обеспечение реактивных свойств нервной ткани (образование рубцов после травмы, участие в реакциях воспаления, в образовании опухолей)
Фагоцитоз (удаление погибших нейронов)
Изоляция синапсов (контактные участки между нейронами)
Источники онтогенетического развития нейроглии:появилась в процессе развития нервной системы из материала нервной трубки.
13. Макроглия (от макро... и греч. glнa - клей), клетки в мозге, заполняющие пространства между нервными клетками - нейронами - и окружающими их капиллярами. М. - основная ткань нейроглии, часто с ней отождествляемая; в отличие от микроглии, имеет общее с нейронами происхождение из нервной трубки . Более крупные клетки М., образующие астроглию и эпендиму, участвуют в деятельности гемато-энцефалического барьера, в реакции нервной ткани на повреждения и инфекции. Более мелкие, так называемые сателлитные клетки нейронов (олигодендроглия), участвуют в образовании миелиновых оболочек отростков нервных клеток - аксонов, обеспечивают нейроны питательными веществами, особенно в период усиленной активности мозга.
14. Эпе́ндима - тонкая эпителиальная мембрана, выстилающая стенки желудочков мозга и спинномозговой канал. Эпендима состоит из эпендимных клеток или эпендимоцитов, относящихся к одному из четырёх типов нейроглии. В эмбриогенезе эпендима образуется из эктодермы.